Корень (в биологии)
КОРЕНЬ в биологии, один из основных вегетативных органов высших растений, возникший в процессе эволюции для закрепления растения в субстрате, поглощения воды и минеральных соединений из почвенных растворов, их преобразования в транспортные формы и перемещения в побеги, синтеза органических веществ (гормоны цитокинин и абсцизовая кислота, ряд алкалоидов и др.), для взаимодействия с почвенными бактериями и грибами. Лишь немногие представители высших растений утратили способность образовывать корни (например, вольфия). Морфологически от побега корень отличается тем, что на нём никогда не формируются листья, а верхушечная (апикальная) меристема всегда прикрыта корневым чехликом. Полагают, что корни возникли из ризомоидов нижнедевонских растений - особых веточек их теломного тела, покрытых одноклеточными нитевидными образованиями (ризоидами), которые росли горизонтально в почве, но убедительного подтверждения этой гипотезы нет.
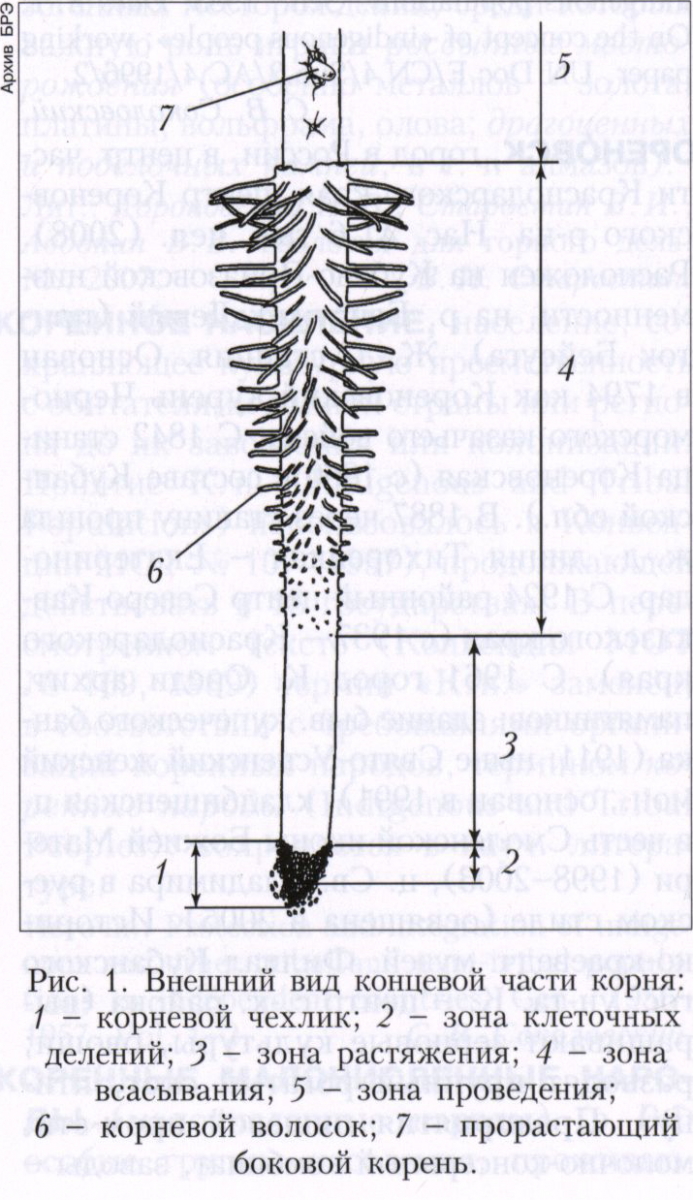 Морфологическое строение корня и корневые системы. Корень имеет цилиндрическую форму и чёткую продольную зональность (рис. 1). На его верхушке (апексе) располагается корневой чехлик, который защищает апикальную меристему и воспринимает направление силы тяжести. Меристема постепенно образует новые клетки чехлика, а краевые его клетки слущиваются и претерпевают апоптоз, вследствие которого выделяются вещества, облегчающие растворение почвенных минералов и служащие питанием или сигналом для почвенной микрофлоры. Прикрытая корневым чехликом апикальная меристема составляет зону клеточных делений. Образовавшиеся клетки усиленно растут в зоне растяжения клеток, что вносит основной вклад в удлинение корня. Суммарная длина обеих зон (иногда их обозначают совокупно как зону роста корня) варьирует в зависимости от интенсивности роста корня и обычно составляет 1-2 мм. В зоне растяжения клеток начинают дифференцироваться первичные постоянные ткани, полный набор которых присущ зоне всасывания, которая обеспечивает поглощение веществ в течение 3-6 суток и преобразуется в зону проведения. Поэтому при росте корня в длину зона проведения удлиняется, а зона всасывания смещается вслед за верхушкой корня в новые участки почвы, сохраняя более или менее постоянную длину (обычно в пределах 0,5-3 см). К молодым участкам зоны проведения приурочено боковое ветвление корней большинства растений.
Морфологическое строение корня и корневые системы. Корень имеет цилиндрическую форму и чёткую продольную зональность (рис. 1). На его верхушке (апексе) располагается корневой чехлик, который защищает апикальную меристему и воспринимает направление силы тяжести. Меристема постепенно образует новые клетки чехлика, а краевые его клетки слущиваются и претерпевают апоптоз, вследствие которого выделяются вещества, облегчающие растворение почвенных минералов и служащие питанием или сигналом для почвенной микрофлоры. Прикрытая корневым чехликом апикальная меристема составляет зону клеточных делений. Образовавшиеся клетки усиленно растут в зоне растяжения клеток, что вносит основной вклад в удлинение корня. Суммарная длина обеих зон (иногда их обозначают совокупно как зону роста корня) варьирует в зависимости от интенсивности роста корня и обычно составляет 1-2 мм. В зоне растяжения клеток начинают дифференцироваться первичные постоянные ткани, полный набор которых присущ зоне всасывания, которая обеспечивает поглощение веществ в течение 3-6 суток и преобразуется в зону проведения. Поэтому при росте корня в длину зона проведения удлиняется, а зона всасывания смещается вслед за верхушкой корня в новые участки почвы, сохраняя более или менее постоянную длину (обычно в пределах 0,5-3 см). К молодым участкам зоны проведения приурочено боковое ветвление корней большинства растений.
Реклама
В зависимости от места образования различают главный корень, формирующийся у зародыша, придаточные корни, отрастающие от стебля и старых частей корни, а при черенковании - и от листа, боковые корни, возникающие в зоне проведения, и верхушечные, развивающиеся при дихотомическом ветвлении корня у плауновидных. Все корни одного растения в совокупности составляют его корневую систему (КС). У хвощевидных, плауновидных, папоротниковидных растений она первично гоморизного типа (рис. 2): состоит из развивающихся на стебле или ризофоре (корневой подставке некоторых плауновидных) придаточных корней, несущих боковые или верхушечные корни; все разветвлённые придаточные корни однотипные. У цветковых и голосеменных растений КС или сложена только системой разветвлённого главного корня, или наряду с ней имеются ветвящиеся придаточные корни. В КС стержневого, или аллоризного, типа главный корень значительно крупнее всех остальных (например, укроп). В КС мочковатого, или вторично гоморизного, типа главный корень не выделяется среди систем мощных придаточных корней или вообще рано отмирает (злаки). Главный и придаточные корни растут по вектору силы тяжести; боковые и верхушечные - под любым углом к нему, что позволяет равномерно заполнять объём почвы. Корни в КС часто дифференцированы на долговечные толстые ростовые (или скелетные) и отрастающие от них тонкие сосущие, существующие всего 2-2,5 месяца. КС проникают в почву на глубину от нескольких мм (центункулус из семейства первоцветных) до 20 м (верблюжья колючка), но у большинства растений основная масса корня располагается до глубины 50-70 см. В стороны корни распространяются в основном на 1-1,5 радиуса проекции кроны, хотя отдельные корни отрастают значительно дальше, а у многих пустынных растений радиус проекции КС в 15-20 раз больше радиуса проекции кроны.
По заполнению объёма почвы выделяют ещё 4 типа КС. Универсальная КС довольно равномерно заполняет объём почвы (присуща мезофитам). Поверхностная КС при небольшой глубине охватывает большую площадь, приспособлена к использованию воды редких дождей и образующейся ночами конденсационной воды (встречается у ксерофитов, в том числе у многих кактусов). Глубинный тип КС обеспечивает проникновение к постоянным грунтовым водам в глубоких слоях субстрата (верблюжья колючка). Комбинированная КС сложена поверхностными и проникающими глубоко в субстрат корня (качим).

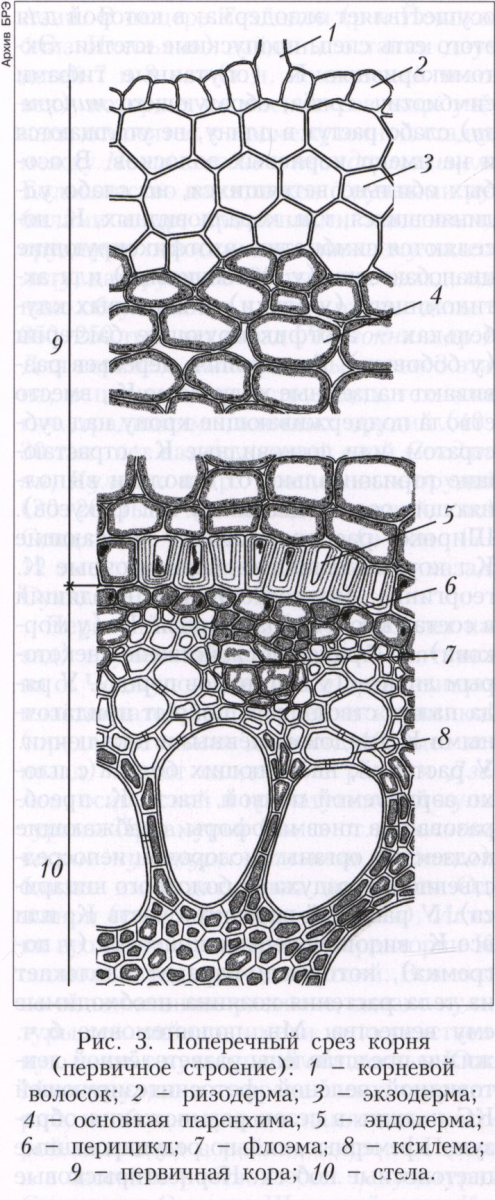
Анатомическое строение корня. Первичная тканевая структура корня формируется в зоне всасывания (рис. 3). Поверхностно в этой зоне располагается слой ризодермы. Клетки последней чаще всего образуют длинные выросты - корневые волоски, которые прорастают в капилляры почвы, что значительно повышает интенсивность поглощения веществ корнем. У водных и околоводных растений корневых волосков обычно нет. В зоне проведения корня ризодерма отмирает и разрушается. Между ризодермой и центральной частью корня (стелой) находится первичная кора. Внешний её слой - экзодерма - прилежит к ризодерме и в зоне всасывания содействует избирательному поглощению веществ, а в зоне проведения становится покровной тканью. Глубже находится паренхима, проводящая поглощённые вещества в стелу и синтезирующая некоторые органические соединения. Внутренний слой первичной коры (эндодерма) участвует в загрузке ксилемы в зоне всасывания и защите продольного транспорта веществ в зоне проведения в стеле. Периферийный слой стелы - перицикл - лишён проводящих тканей и представлен обычно одним слоем паренхимы. В зоне проведения в перицикле происходит локальная дедифференциация паренхимных клеток с образованием апикальных меристем боковых корней, впоследствии прорастающих сквозь первичную кору материнского корня в почву. Внутрь от перицикла находится радиальный проводящий пучок: его ксилема имеет вид ребристого цилиндра, в желобках между рёбрами которого проходят изолированные друг от друга тяжи флоэмы. Для однодольных характерны многорёберные пучки, для остальных групп растений - малорёберные. Ксилема радиального пучка содержит наряду с водопроводящими элементами и паренхимные клетки, которые активно (при затрате энергии) выделяют в просветы водопроводящих элементов растворимые в воде низкомолекулярные вещества и ионы. Вслед за ними в водопроводящие элементы из паренхимных клеток начинает поступать вода и возникает избыточное давление, обычно составляющее 1-2 атмосферы (иногда достигает 6 атмосфер). Корневое давление обусловливает продвижение воды по ксилеме корня к его основанию и далее в побеги. До распускания листьев корневое давление полностью обеспечивает водоснабжение побегов растения, а затем его роль значительно снижается и оно замещается сосущей силой транспирирующих листьев.
 У двудольных и голосеменных растений уже у границы между зонами всасывания и проведения начинается вторичное утолщение корня. Перицикл становится многослойным из-за пролиферации паренхимы, по его наружному краю формируется перидерма, вследствие чего первичная кора целиком отмирает и разрушается. В стеле дифференцируется непрерывный слой камбия, одни участки которого возникают в радиальном пучке между флоэмой и ксилемой, а другие - в перицикле, снаружи от рёбер ксилемного цилиндра. Перициклические участки камбия откладывают только паренхиму первичных лучей. Пучковые участки камбия откладывают внутрь вторичную ксилему и наружу вторичную флоэму. У древесных растений со временем в корне между первичными лучами образуются и вторичные.
У двудольных и голосеменных растений уже у границы между зонами всасывания и проведения начинается вторичное утолщение корня. Перицикл становится многослойным из-за пролиферации паренхимы, по его наружному краю формируется перидерма, вследствие чего первичная кора целиком отмирает и разрушается. В стеле дифференцируется непрерывный слой камбия, одни участки которого возникают в радиальном пучке между флоэмой и ксилемой, а другие - в перицикле, снаружи от рёбер ксилемного цилиндра. Перициклические участки камбия откладывают только паренхиму первичных лучей. Пучковые участки камбия откладывают внутрь вторичную ксилему и наружу вторичную флоэму. У древесных растений со временем в корне между первичными лучами образуются и вторичные.
Специализация и метаморфозы корней. В ходе эволюции корни многих растений приобретали дополнительные функции, более или менее глубоко изменяя свою структуру. У эпифитных орхидей в первичной коре корня имеется фотосинтезирующая ткань - хлоренхима, а вместо ризодермы развита ткань, состоящая из мёртвых клеток (веламен), неизбирательно впитывающая стекающую по растению дождевую воду; избирательное поглощение осуществляет экзодерма, в которой для этого есть специальные пропускные клетки. Эктомикоризные корни (опутанные гифами симбиотического гриба, образующего микоризу) слабо растут в длину, не утолщаются и не имеют корневых волосков. В особых обильно ветвящихся, но слабо удлиняющихся, так называемых коралловидных, корнях поселяются симбиотические азотфиксирующие цианобактерии (у саговниковых) или актиномицеты (у ольхи); в корневых клубеньках - азотфиксирующие бактерии (у бобовых). Ряд тропических деревьев развивают надземные ходульные корни, вместо ствола поддерживающие крону над субстратом, или досковидные корни, отрастающие горизонтально от ствола и выполняющие роль подпорки (у многих фикусов). Широко распространены запасающие корни: корневые шишки (придаточные корни георгины, аспарагуса) или входящий в состав корнеплода главный корень (у моркови). Корни-прицепки свойственны некоторым лианам (у отдельных видов перца). У ряда пальм ствол густо покрыт придаточными корнями, видоизменёнными в колючки. У растений, населяющих болота с плохо аэрируемой почвой, часть корня преобразована в пневматофоры, снабжающие подземные органы кислородом непосредственно из воздуха (у болотного кипариса). У растений-паразитов часть корня или все корни видоизменены в присоски (у погремка), которыми паразит извлекает из тела растения-хозяина необходимые ему вещества. Многие подостемовые большую часть жизни представлены разветвлённой лентовидной зелёной фотосинтезирующей КС и лишь в сезон размножения образуют эфемерные, сильно редуцированные цветоносные побеги. Корнеотпрысковые растения размножаются корневыми отпрысками, возникающими в перицикле корня.
Корни многих растений используют как пищевое, лекарственное и техническое сырьё.
Лит.: Раздорский В. Ф. Анатомия растений. М., 1949; Федоров А. А., Кирпичников М. Э., Артюшенко 3. Т. Атлас по описательной морфологии высших растений. Стебель и корень. М.; Л., 1962; Воронин Н. С. Эволюция первичных структур в корнях растений // Ученые записки Калужского педагогического института. 1964. Вып. 13; Лотова Л. И. Морфология и анатомия высших растений. М., 2001; Ботаника: на основе учебника Э. Страсбургера [и др.]. М., 2007. Т. 1-2; Ботаника. М., 2007. Т. 3: Тимонин А. К. Высшие растения; Физиология растений / Под редакцией И. П. Ермакова. 2-е изд. М., 2007.
А. К. Тимонин.